Influence of microorganisms
MENU
Home
Categories
Classification
Bacteria
About Microscopic Organism
Microorganism
Definition
Function
Microbes
Types
Virus
Contact
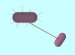
Structure and Function of Bacterial cell Component
About Microscopic Organism
/
Source:
RELATED POSTS
- Composition of Bacterial cell wall
- Structure and Function of Cytoplasm
- Function of pili in Bacterial cell
- Function of Bacterial plasmid
LATEST POSTS
Functions of Pili in Bacteria
June 12, 2018
Protein Capsule Bacteria
December 5, 2018
Prokaryotes, Eukaryotes, and Viruses
February 15, 2018
Bacterial Infections
October 21, 2018
ADS BY GOOGLE
All you need is here
CATEGORIES
Classification
Bacteria
About Microscopic Organism
Microorganism
Definition
Function
Microbes
Types
Virus